ИЗУМИТЕЛЬНАЯ
АСИММЕТРИЯ
Ю.
Чайковский
Живая природа оказалось очень трудным
орешком для теоретиков науки, преподнося им
все новые неожиданности на пути познания. О
причинах такой каверзности говорил в
статье «Теоретическая биология? Ее все еще
нет…» В. В. Налимов, доктор технических наук
(№ 7, 1979 год). На то же сетовали биолог М. В.
Минина и математик А. М. Молчанов («Три витка
в спирали спорта», № 9, 1979 год), обсуждая
препятствия, стоящие на пути теоретизации
биологии, на том пути, которые иные науки
уже успешно проходят. В центре всех
дискуссий о теоретической биологии всегда
стоит вопрос: отличается ли живая природа
от неживой столь существенно, что «ей нужны
свои законы», или просто наука пока не
подобрала нужный ключ из имеющихся в ее
запасах? Вопрос этот крайне важен для
познания вообще. Исследователи
предпринимают различные попытки для его
решения.
Предлагаемая статья – одна из таких
попыток. Автор ее рассказывает о
неожиданностях, с которыми встретились
математики, подвергнувшие теоретическому
рассмотрению закономерности разнообразия
организмов.
Листочки
с одной ивы
«Я до сих пор живо, помню, как однажды, когда
я еще был ребенком, отец привел меня на край
города, где на берегу росли ивы, и велел мне
сорвать наугад сотню ивовых листочков…
Вернувшись домой, мы расположили их в ряд по
росту, как солдат. Затем отец через кончики
листьев провел кривую и сказал6 «Это и есть
кривая Кетле. Глядя на нее, ты видишь, что
посредственности всегда составляют
подавляющее большинство и лишь немногие
подымаются выше или так и остаются внизу».
Так писал известный современный математик
Б. ван дер Варден в своем учебнике
математической системы.
Пример Кетле действительно поучителен: эту
кривую можно получить, не только, ранжируя
листья или солдат, но самыми неожиданными
способами; дело в том, что кривая Кетле – не
что иное, как гауссово распределение,
называемое еще нормальным распределением
случайных величин.
Почему гауссово распределение называют
нормальным? Прежде всего потому, что
результат почти всякого массового
измерения однотипных величин дает одну и ту
же картину: отдельные измерения
отклоняются от своего среднего («нормы») с
частотой, близкой к f(x). Сам Гаусс, еще до
работ Кетле, обнаружил этот факт для таблиц
координат звезд, но функцию f (x) теория
вероятностей знала и за сто лет до Гаусса.
Так при чем же здесь Кетле и чем вообще это
нам интересно? Дело в том, что именно Адольф
Кетле, бельгийский математик и статистик,
заложил то воззрение на мир чисел, которое
лежит в основе основ современного
естествознания и в котором уже поэтому
полезно время от времени сомневаться.
Вот что писал Кетле: «Все элементы
организмов колеблются около среднего
состояния, и … изменения, происходящие под
влиянием случайных причин, подчинены такой
точности и гармонии, что их все можно
перечислить наперед». Другими словами,
можно, например, зная средний рост мужчины а
и дисперсию роста (грубо говоря, среднее
отклонение от этой величины) вычислить,
сколько какого обмундирования потребуется
для дивизии. Сейчас это настолько очевидно,
что трудно осознать, почему книгами Кетле
зачитывались многие современники
Белинского и Гоголя, но все-таки
постараемся.
Статистика как способ описания массовых
явлений известна с шекспировских времен, но
до Кетле она в основном сводилась к
простейшим подсчетам, и недаром ее называли
«политической арифметикой» - она не шла
далее суммирования величин и их деления.
Насколько она была еще беспомощна, видно из
такого примера. В учебнике 1937 года читаем:
гигантом следует считать человека, чей рост
ни у одного народа нельзя считать
нормальным; а поскольку есть данные, что на
Огненной Земле нормален рост до 7 футов, то
есть до сажени (213 сантиметров), то гигант –
тот, кто выше 7 футов. Итак, рекомендация (важная,
например, для рекрутских наборов) зависела
от того, какие слухи дошли до моряков, а не
от того, что уже двухметровый человек
торчал из тогдашней толпы по шею.
Статистическая логика вошла в описательные
науки только с Кетле, и тогда открылась
возможность достаточно точных прогнозов,
не всегда очевидных с первого взгляда. Так
нетрудно вычислить средний рост и
дисперсию роста; зная их можно предсказать,
что приблизительно каждый сорок четвертый
солдат окажется ниже среднего роста более
чем на удвоенную дисперсию, зато каждый
сорок четвертый будет на такую же величину
выше – все симметрично. Эти сильные и в то
же время редкие отклонения можно назвать
нетипичными, откуда и рабочее понятие –
типично (и в этом смысле нормально), то, что
близко к среднему.(Удобство понятие,
впрочем, бывает обманчиво, как мы вскоре и
увидим.) Идя этим путем, Кетле сформулировал
идею социального благополучия как баланса
средних значений и идею эволюции общества
как направленного сдвига этих средних.
Свою первую книгу он издал в 1835 году, а два
года спустя молодой Дарвин записал в
блокнот свои первые мысли об эволюции
организмов. Дарвин и Кетле – оба они
выражали идею своего века: знание как
статистику, мир как баланс средних, а
эволюцию – как сдвиг средних, (Кстати, они
имели и общего предшественника – юриста и
ботаника Альфонса Декандоля; Кетле
использовал его статистические данные по
преступности, а Дарвин – по географии
растений, но первый делал акцент на
устойчивость средних, а второй – на их
сдвиг.)
Статистика
отбора
Как не далек был Дарвин от статистиков, но
уже в его первых записях мелькает: «
уравновешенное число форм», «каждое
поколение должно иметь столько же
продолжающих существовать, как и сейчас», «строгая
постепенность перехода», « закон малых
различий, производящих более плодовитое
потомство», «вид, подобно погоде, на
протяжении длительного отрезка времени
остается в среднем относительно
однообразным» и т. п. Подобен
статистический элемент и в «Происхождении
видов», а позже, в 1873 году, Дарвин писал
прямо: «По исследованиям Кетле… люди в
отношении роста могут быть сгруппированы
вокруг средней величины. …Мы можем
предположить, что таков обычный закон
изменчивости у всех органов каждого вида…
что при неблагоприятных для вида условиях…
группировка уже не была бы симметричной по
отношению к средней величине
рассматриваемого органа. В этом случае… с
течением времени выжили бы только те особи,
у которых такие органы имели бы надлежащую
величину». Другими словами, отклонение
распределения от кривой Гаусса – вот
первый шажок эволюции.
Эта мысль стала главной, когда в ХХ веке
дарвинизм стал вооружаться статистикой.
Если условия среды требуют, чтобы какой-то
признак организма стал более определенно
выраженным или просто увеличился, то
материал для этого найдется: его представят
те особи, которые, отклоняясь от «посредственностей»
в нужную сторону, производят тем самым
больше потомства. Гауссова кривая как бы
вытянет «голову» и подожмет «хвост», чем и
сдвинет среднее; в следующих поколениях эта
смешанная кривая плотности станет основой
для новых сдвигов – вот и элемент эволюции.
Не вступая в дискуссию о работоспособности
самого этого механизма, зададим себе
примитивный вопрос: а всегда ли у плотности
распределения искомого признака есть «хвост»,
который может быть поджат, и «голова»,
которая ползет куда надо? Например, если
какой-то вид рыб надо поджимать, чтобы он
начал заботиться?
Явление
героини
Мы уже видели, как гауссово распределение
симметрично: всякому уклонению от нормы
здесь можно поставить в соответствие
противоположное (короткому листу – длинный,
яркому таланту – бездарность и т. д.). Нашей
же героиней станет асимметрия, причем нас
будет интересовать не такая мелочь как
неравенство правого и левого «хвостов» на
кривой, а радикальная асимметрия, когда
одного из «хвостов2 нет и в помине.
Парадокс состоит в том, что Варден-отец был
не вполне прав, давая урок будущему
статистику: посредственность далеко не
всегда заключена между талантом и
бездарностью, если понимать эти термины как
статистические. Разумеется, если в вузе
подвергнуть всех студентов какому-нибудь
тесту на умственное развитие, то результат
прекрасно ляжет на гауссово распределение,
но это потому, что тест специально на это
рассчитан (и, возможно, только на это). С теми
же студентами можно проделать и другой
эксперимент, который даст абсолютно другой
результат: надо подождать лет тридцать –
сорок, а затем подсчитать, сколько кто
написал научных работ. Вопреки ожиданиям,
распределение работников науки по числу
написанных ими статей имеет однохвостую
плотность: около трети не опубликовало ни
одной статьи, столько же имеет по одной-две
публикации; и далее – чем более статей, тем
меньше число авторов им обладает. Самое же
парадоксальное то, что большая часть всех
статей обязательно окажется написанной
теми немногими, кто имеет по тридцать
статей и более. Если рассуждать по Вардену-отцу,
пришлось бы признать, что почти все «остались
внизу», а «посредственностей» (то есть
авторов, имеющих по 5 –6 статей – близкие к
среднему результату печатной деятельности)
мало.
Точно так же распределяются и сами статьи
по их цитируемости: около трети не
цитированы никем и никогда, столько уже
упомянуты по 1 –2 раза, а основная масса
ссылок приходится на малую длю статей.
Аналогично получается с выживанием
организмов и видов. Проще всего убедиться в
этом на примере колонии бактерий, хотя
интересующее нас обстоятельство типично
для всех организмов.
Пусть каждая бактерия делится ежечасно,,
причем все они выживают. Тогда через час из
одной бактерии родится две, через два часа -
четыре, через три – восемь, а через сутки –
почти 17 миллионов. Рано или поздно их станет
столько, что они не смогут пропитаться, и
тогда-то начнется самое интересное. Ведь
если колония сохраняет свой общий размер,
соответствующий поступлению пищи извне, то
в среднем на одну бактерию будет
приходиться не два потока, а только один.
Выберем одну бактерию и будем следить за ее
потомством, затем проделаем то же с другой
бактерией той же исходной колонии и так
далее. Потомство одной особи, полученный
бесполым путем, называем клоном; если
следить за десятком тысяч таких кланов, то
окажется, что через 100 поколений
сохранилось всего около 200 кланов, тогда как
остальные 9800 за это время вымерли. Зато
оставшиеся в живых 200 клонов окажутся
весьма различными по численности: в них
будет в среднем по 50 бактерий на клон, что и
даст необходимые 10000 особей (неизменный
баланс – одну особь на исходный клон).
Как мы видим, распределение живых клонов по
численности тоже оказывается «однохвостым»;
при этом клонов, представленных одной-единственной
особью, то есть средним числом, будет всего
три-четыре (из десяти тысяч!), следовательно,
термин «среднее» не несет здесь того смысла
«наиболее вероятного», что в гауссовой
статистике. Наоборот – наиболее вероятная
численность здесь всегда далека от средней:
она или гораздо ниже (в нашем случае –
нулевая численность вымерших клонов), или
гораздо выше.
Заметим, что приведенные цифры – результат
чисто вероятностного расчета (подробно
проведенного, кстати, в № 8 журнала «Генетика»
за 1977 год): все особи приняты одинаковыми,
следовательно, в их вымирании не было
никакого отбора лучших, а одна только
чистая случайность.
К «однохвостым» плотностям бесполезно
подходить с обычной статистической меркой:
среднее значение здесь не имеет особого
смысла, зато термин «самое вероятное» имеет
сразу два противоположных смысла – так,
самая вероятная численность выжившего
клона равна единице, но наугад взятая
бактерия будет, вероятнее всего,
принадлежать не такому клону, клону
численностью более пятидесяти. Если в
обычной гауссовой статистике можно хоть в
каком-то смысле говорить о норме как о
среднем, то здесь норму одним числом
выразить нельзя, а приходится вводить новый
язык, язык кластеров.
Кластер – это компактное множество
элементов, имеющих близкие свойства, причем
каждый кластер имеет свои характеристики,
свою норму. Так, разумно сравнивать размеры
«пустых» кластеров (доли непечатающихся,
нецитированных, вымерших и т. д.) или размеры
«ведущих» кластеров (доли авторов, пишущих
половину всех статей, доли клонов,
составляющих треть всей колонии и т. д.).
Некоторые начальники, не зная свойств «однохвостой»
статистики, считают своим долгом подымать
публикуемость – свою и своих сотрудников,
но однобокость, естественно, только
перемещается: вымученные публикации
попадают в кластер нецитируемых. Стали
поговаривать, что работников надо
оценивать по их цитируемости, и результат
нетрудно предсказать: возрастет кластер
авторов, патологически цитирующих друг от
друга, но не цитируемых никем более. А ведь
работники с низкой публикуемостью –
нормальный кластер работоспособного
коллектива, и подавлять этот кластер –
значит подрывать работу коллектива: на них
обычно ложится основной груз промежуточных
экспериментов, реферирования и внутреннего
рецензирования, не говоря уж про
материальную базу лабораторий.
В чем же причина того, что одни случайные
процессы укладываются в симметричные
плотности типа гауссовой, а другие – «однохвосты»?
Мы живем в мире случайностей, и они все
время накладываются одна на другую. Если
независимые случайности просто
складываются, а мы измеряем эту сумму, то
результаты наших измерений почти всегда
уложатся в гауссово распределение (в этом
заключается «центральная предельная
теории вероятностей). Сходная ситуация
получится, если случайности перемножаются,
так как при этом складываются их логарифмы:
кривая перекосится, но хвостов у нее
останется два, как и у кривой.
Однако взаимодействие случайностей может
быть гораздо сложнее, нежели сложение и
перемножение: так, в примере с бактериями
случайное выживание порождает возможность
следующей случайности (выживания в
следующем поколении), тогда как случайная
гибель никакой новой случайности породить
не может – вот, наглядно говоря, и источник
асимметрии, порождающий однохвостность. В
примере с публикациями он не так очевиден,
но таков же в сущности: вероятность
следующей статьи растет с ростом числа уже
опубликованных данным автором работ - ему
легче и писать, и печататься.
«…Свой
подвиг бесполезный»
Итак, дарвиновское «уравновешенное число
форм» скрывает в себе никак не колебания
вокруг среднего, а бурю, губящую почти всех
и дающую право на колоссальное потомство
немногим. Это повторим, не имеет пока
никакого отношения к отбору (к «закону
малых различий, производящих более
плодовитое потомство»), так как
плодовитость всех особей предполагалось в
наших расчетах одинаковой. Что же будет,
если в этой буре примут удары не равные, а
несколько различные по плодовитости
организмы? Другими словами, что будет, если
в наших выкладках учесть влияние
естественного отбора? Оказывается, нового
появится немного: процесс, в котором
случайности порождают друг друга, слишком
хаотичен, чтобы чувствовать малые вариации
плодовитостей.
Пусть мы следим за клоном, каждая особь
которого оставляет в среднем 1,01 потомка, то
есть с вероятностью 0,495 гибнет, а с
вероятностью 0,505 делится на две. Легко
вычислить, что такой клон со вполне
ощутимый вероятностью 1/50 доживает до
бесконечности, тогда как любой клон,
оставляющий в среднем на особь одного
потомка или менее, наверняка рано или
поздно вымрет. Еще в 1922 году Рональд Фишер,
крупный английский математик и
основоположник эволюционной статистики,
получив подобный результат, счел его
решающим аргументом в пользу нового
дарвинизма, ведь если полезная мутация
возникает хотя бы сто раз, то ее обладатели
наверняка сохранятся и в конце концов
вытеснят всех остальных. Он, повидимому,
просто не подумал о самых очевидных
трудностях – ведь отбору подвергается не
мутации, а их полезные комбинации (их не то
что сто раз, а и дважды не всегда дождешься),
причем отбор каждого полезного клона
должен закончится за ограниченное время. В
течение этого времени не только может
сохраняться любой клон-конкурент, но и
будет неизбежно меняться сами условия
конкуренции. Другими словами, стоит немного
приблизить расчет к реальности, чтобы
получились противоположные выводы.
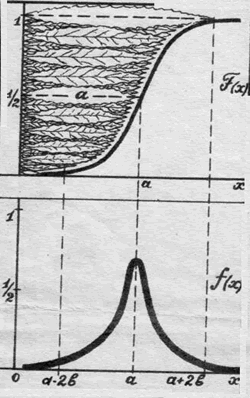 |
Общеизвестное
нормальное (или гауссово)
распределение - плотность кривой Кетле:
если кривая Кетле - результат простого
ранжирования листочков, то кривая
Гаусса показывает, какова доля
листочков, имеющих одинаковую длину.
Буквой "а" обозначена средняя
величина (в нашем примере - средняя
длина листочков ивы), а буквой "в" -
дисперсия этой величины, то есть
характеристика степени отклонения
случайных значений (случайных длин
листочков) от средней величины "а".
|
Прежде всего, отбор одной мутации не должен
занимать больше нескольких сотен поколений:
во-первых, этого требует реальные
наблюдаемые темпы эволюции, а во-вторых,
редко в какой среде плодовитость в 1.01 может
удержаться сто поколений подряд (для
фишеровских расчетов нужно именно подряд).
Далее, в самом ли деле полезная мутация
вытесняет все вредные с течением времени?
Не окажется ли, что новые мутации возникают
ранее, чем отбор успевает решать судьбу
прежних?
Увы, расчет, проведенный для конечных
времен, показывает именно последнее:
полезный клон растет (при плодовитости, не
намного превышающей единицу) настолько
хаотически, а вредные клоны поддерживают за
счет новых мутаций свою общую численность
так регулярно, что победа полезного клона
редко возможна. Если мутанты действительно
различаются по плодовитости только на
несколько процентов, то состав большой
колонии будет определяться законами
мутагенеза, а никак не отбора. Если
допустить, как мы это видим в природе, что на
одну полезную мутацию приходится тысяча
вредных и что мутации возникают во всех
поколениях, а не только, как у Фишера, в
начальном, то получим, что общая
численность вредных мутантов обычно
превосходит численность полезных. Фишер
был прав, утверждая, что одна полезная
мутация может вытеснить одну вредную, но
оказывается, что редкие слабовыгодные
мутации тонут в потоке слабовредных. Отбор,
как многократно проверенно и в природе, и в
лаборатории, и в практике селекционеров,
эффективно действует тогда, когда разница в
плодовитости участников велика _ когда один
тип организмов размножается свободно, а его
конкуренты почти не размножаются.
Разумеется. малая разница в плодовитости
может помочь выживанию, но она не
обеспечивает вытеснения конкурентов.
Прочтя это, читатель, по-видимому готов
спросить: неужели эти простые расчеты не
были известны тем, кто пятьдесят лет назад
создал статистическую концепцию отбора?
Ответить трудно – и были бы и не были. Дело
ведь не столько в фактах, сколько в желании
их видеть. Еще при Дарвине некоторые
сомневались что отбор может преодолевать
бурю случайного истребления. Учение
Дарвина вызвало бурные споры, в частности
баталию осенью 1870 года на страницах журнала
«Nature» о количественной стороне проблемы
отбора. Позже, в декабре, на спор
откликнулся энтомолог Сэмюэл Скеддер из
Египта, писавший в журнал: можно ли говорит
о подборе качеств взрослой бабочки. если до
этой стадии доживает малая доля куколок, до
куколки – малая доля гусениц, а до гусеницы
– малая доля яиц? Скеддера пожало: гусеница
гротескной формы и угрожающей окраски
буквально кишит пожирающими ее паразитами,
тогда как близкий вид гусениц для них
несъедобен. Почему отбор растрачивал силы
на такие детали, как рисунок гусеницы или
взрослой бабочки, а не на борьбу с
паразитами?
Скеддер не отрицал сразу роль отбора. он
предлагал сперва выяснить, действительно
ли яркая окраска существенно улучшает
судьбу бабочек, но его оппоненты стали
обсуждать не это. Излагая свои наблюдения,
Скеддер не избежал мелких ошибок, и
дискуссия, как водится, закрутилась вокруг
них, а суть дела осталось без внимания.
Правда, сам Дарвин вставил в тест
последнего издания «Происхождение видов»
абзац, где разбираемая нами трудность
отмечена. Признав, что основная масса яиц (да
и некоторых взрослых организмов)
истребляется случайно, без связи с отбором,
он закончил оптимистически: «и тем не менее
из числа тех, которые выживут, наиболее
приспособленные особи… будут размножаться
в большем числе». Насколько «в большем» (можно
ли эту разницу заметить за все время
существования вида?), Дарвин не сказал,
Вскоре возражение Скеддера забыли, и только
на век принес наконец остроумный ответ на
него: можно, оказывается, повальное
выедание яиц и молоди назвать особым видом
отбора – «неспецифическим» и считать, что
он приводит просто к росту плодовитости.
Опыт, как видим, и ставить незачем. Тем более
незачем выяснять, на каких постулатах
покоятся формулы, пока они дают то, чего от
них хотели. (Порадоксально: самые грубые
расчеты Фишера в течение полувека казались
биологам вполне убедительными, когда же
японский исследователь Моту Кимура уже в
наши дни уточнил их и получил вывод о
неэффективности отбора слабых вариаций, то
его все наперебой упрекают в биологической
некорректности его формул).
Так как же, имеет ли отбор слово в судьбе
организмов? Прежде всего, свое слово, пусть
и самое веское, он может произнести только
тогда, когда случайность «предоставит ему
слово», - полезный мутант должен случайно
пережить первые поколения, чтобы,
размножившись, иметь возможность
продемонстрировать свою жизнеспособность
в различных ситуациях. Далее, его слово
веско никак не в отношении мелких вариаций.
Следовательно, если процесс изменчивости
будет предъявлять на суд отбора достаточно
крупные преимущества и притом не слишком
редко, то отбор, безусловно, сможет вести
эволюцию. Однако многие еще считают, что
важные для эволюции полезные уклонения
мелки и проявляют пользу только «в среднем»;
они уверены, вслед за статистиками прошлого
века, что сдвиг средних – это и есть
эволюция, но теперь-то уже известно, что
эволюция – не сдвиг средних, а смена
кластеров. Природу же, основанную на
сдвигах средних значений, лучше всего
характеризовать словами Тютчева:
|
Поочередно
всех своих детей,
Свершающих свой подвиг бесполезный,
Она равно приветствует
своей
Всепоглощающей и миротворной бездной.
Мутации
и гиперболы. |
Не будем, однако сразу впадать в уныние:
пессимистический вывод предыдущей главы
основан, к счастью, не на реальных фактах
природы, а на допущениях, плохо продуманных
авторами двадцатых – тридцатых годов
нашего века. Ведь даже у Дарвина природа -
далеко не такое болото мелких вариаций.
Слов нет, «закон малых различий,
производящих более плодовитое потомство»,
сформулирован именно Дарвином и защищался
им более сорока лет, но фактическая
аргументация Дарвина почти не затрагивала
этот закон. Сравнивая различные
приспособления, Дарвин чаще обращался к
различию родов или видов, а не к мелким
вариациям (о которых он обычно вспоминал,
доказывая всеобщий характер изменчивости),
и дарвинизм не сильно пострадает, если
отвергнуть их роль в происхождении
приспособлений. Такую модификацию
дарвинизма уже предлагал Гуго де Фриз,
знаменитый ботаник генетик, в 1903 году: на
ряде примеров он доказывал, что вид
происходит в результате «видовой мутации»,
то есть одним скачком, а не путем накопления
мелких вариаций, роль же отбора он видел
только в изъятии неудачных видов. После
короткого периода популярности учение де
Фриза сошло со сцены, когда удалось
показать, что его «видовые мутации» – не
совсем то, что называли мутациями, да и
порождают не совсем то, что называли видами.
Опять, как и в случае со Скеддером, критики,
придравшись к фактическим неточностям, не
обсудили суть дела.
Не вдаваясь в старую дискуссию о том, в чем
де Фриз как эволюционист был прав, а в чем
нет, вспомним лучше об одной эволюционно-статистической
работе, стимулированной учением де Фриза. В
этой работе происхождение видов впервые
было рассмотрено как феномен «однохвостой»
статистики.
Джон Кристофер Виллис, как почти все
английские ботаники, был в юности очарован
учением Дарвина, однако, видя его слабую
фактическую обоснованность, решил собрать
недостающую аргументацию сам. Однако в
отличие от большинства он не задался целью
подтвердить (или опровергнуть) дарвинизм, а
решил взглянуть на природу «подобно
инопланетянину, цивилизация которого еще
только стоит на пороге рождения науки». Это
ему в общем удалось, возможно, потому, что он
долгие годы работал на Цейлоне и в Бразилии,
вдалеке от эволюционных школ. Уже на склоне
лет, будучи членом Королевского общества и
автором фундаментального «Словаря
цветковых растений и папоротников», он взял
подсчитать, как распределены виды по родам,
а роды – по семействам. Тут он и обнаружил
уже известный нам феномен однобокости.
В своем «Словаре» он насчитал 12571 род
цветковых, из которых 4853 рода содержали по
одному виду – поразительно! Ведь понятие
рода для того и введено, чтобы объединять
сходные виды, на одновидовой род принято
смотреть как на исключение из правила – то
ли следствие плохой работы систематика, то
ли следствие вымирания видов. Но вот
выясняется, что таких родов – более трети, а
вместе с двух-видовыми - более половины всех
родов цветковых. Для сравнения Виллис
просчитал некоторые семейства низших
растений, а также жуков, змей, ящериц и всюду
нашел ту же закономерность. Разные люди,
разная изученность и разный возраст
таксонов, а результат один и тот же.
Виллис построил графики: по оси абсцисс
число видов в роде. а по оси ординат
количество соответствующих родов – и
получил хорошие гиперболы. Когда же графики
увидел приятель Виллиса - математик Гаролд
Энди Юл - и посоветовал откладывать по осям
графика не сами величины, а их логарифмы, то
изумление ботаника еще более возросло:
почти все точки аккуратно легли на прямую.
Более того, прямые для разных семейств
легли почти параллельно.
Виллис понял, что открыл закон системы
организмов, не связанный прямо с учением
Дарвина. Действительно, если бы
разнообразию условий обитания, как учил
Дарвин, то оно не могло бы так четко
следовать закону, игнорирующему всякую
экологию. Гиперболы подтверждали, как счел
Виллис, правоту де Фриза: структура рода
отражает еще не понятый закон мутаций. Это
допущение вскоре получило веское
подтверждение.
Книга Виллиса вышла в 1922 году, в тот же год,
когда Фишер опубликовал свою первую статью
по динамике выживания клонов. Теперь
настало время основательно потрудиться Юлу:
он взялся объяснить только что открытый
Виллисом закон математически и
воспользовался тем же аппаратом, что и
Фишер, - теорией ветвящихся случайных
процессов. Юл положил, что вид делится на
два скачком (то есть в результате «видовой
мутации»), и получил однохвостое
распределение видов по ролам.
Далее Юл допустил, что роды образуются
подобно видам – за счет «родовых мутаций»,
и после этого получил кривые распределения
видов по родам, почти не отличимые от
гипербол Виллиса.
Совпадение оказалось так точно, что удивило
самого Юла (ведь в группировке таксонов
некоторый произвол систематиков все-таки
неизбежен). Для нас, однако. самое
интересное здесь то, что для совпадения
наблюдаемых кривых с математическими
требовалось брать вполне определенные
соотношения частот «видовых» и «родовых»
мутаций, причем оказалось, что скорость
родообразования мало отличается от
скорости видообразования.
И еще: каждый вид в модели Юла мог порождать
новые независимо от своего возраста и
численности, что было против всех
представлений об изменчивости – ведь
мутация мыслится как нечто, происходящее в
организме, то есть тем более вероятное, чем
больше самих организмов. Это было уже
серьезным вызовом господствовавшим
эволюционным представлениям: ведь, по
Дарвину, новый род образуется за счет
многократных видообразований, а не за счет
однократной «родовой мутации», почти столь
же частой, как и «видовая мутация».
Абсурдные постулаты? Допусти, но как быть с
фактом точного совпадения кривых? Разве в
основе наук не лежит тезис: практика –
критерий истины? Так или иначе, это было
явно нечто новое в эволюционизме. До теории
в нынешнем смысле было еще, разумеется,
далеко, но не будем слишком строги.
Где же истина?
Именно тогда, в двадцатые
годы, в биологию вошел и утвердился тот
тезис (заимствованный из физики и химии),
что истина проверяется только
экспериментом. Четкие повторяющиеся
результаты — вот истина, а отвлеченные
рассуждения — вообще не наука. Так судили
тогда даже ведущие биологи. Однако
крайности обычно приводят к абсурду, и один
из них — судьба идеи Виллиса — Юла: те
предъявили четкий, многократно проверенный
результат, но ученый мир пренебрег и самими
наблюдениями, и фактом многократного
точного совладения этих наблюдений с
простой и ясной теорией. Предпочтение было
отдано чистому умозрению фишеровской школы,
потому что оно было приемлемее для
тогдашнего «рационального» мировоззрения.
Впрочем, сама регулярность кривых
требовала от систематиков какой-то реакции,
и она последовала: кривые Виллиса не
представляют, оказывается, интереса для
систематики, поскольку таким же кривым
подчиняется распределение частот букв в
языке, химических элементов в земной коре,
ученых по их публикациям и многое другое.
Сходство кривых чисто случайно, поскольку
эти явления несопоставимы, они
определяются разными законами природы —
таков был приговор тех, кто считал себя
специалистами в этой, еще не родившейся,
дисциплине. (Оно действительно случайно — в
том смысле, что все непознанное называют
случайным).
Отчасти была виновата
математика, которой пользовался Юл,—
теория ветвящихся процессов только
зарождалась. Юлу пришлось почти все
доказывать самому, и он не мог не ввести
ряда упрощений, ставших вызовом биологии.
Сейчас изучены ветвящиеся процессы самых
разных типов, и Юл мог бы добиться
совпадения с виллисовыми кривыми, не
шокируя биологов. Однако одно осталось бы:
близкие скорости видо- и родо-образоаания.
Основная же беда была в
неподготовленности почвы: во все времена
новые законы природы воспринимались с
трудом, а Виллис и Юл указали на один из
самых общих законов природы — "однохвостую"
статистику. Разумеется, и в те дни не стоило
бы говорить, что разные законы природы
несопоставимы: разве и тогда не было
принято сопоставлять колебания маятника с
электромагнитными? Однако только в наше
время, с развитием теории систем,
становится ясно, чего именно не поняли
критики...
(статья оборвана)